Nasi-García, Luca¹ ²; Jiménez-González, Sonia¹ ² *
¹ Facultad de Medicina, Universidad de Granada (UGR)
² Academia de Alumnos Internos, Facultad de Medicina, Universidad de Granada
* Autor de correspondencia: jimenezglezsonia@gmail.com
Resumen
Los astrocitos son las celulas más abundantes en el sistema nervioso central (SNC) humano. Tradicionalmente, la investigación relacionada con los astrocitos se ha basado en dos premisas basicas: que los astrocitos forman un sincitio homogéneo en todo el SNC y que sus funciones son meramente de soporte y homeostáticas. La ruptura con estas premisas ha dado lugar a un nuevo paradigma en la investigacion relacionada con los astrocitos, en el que cobra relevancia una red funcional de astrocitos vinculados al procesamiento de la informacion y al comportamiento. Utilizando la definición de “conjunto funcional astrocítico” de Delgado et al., buscamos estas estructuras en todo el sistema nervioso central, describiendo conjuntos funcionales astrocíticos definidos y subconjuntos astrocíticos que tambien podrían definirse como conjuntos funcionales astrocíticos.
Palabras clave: Astrocitos, conjuntos funcionales, cojuntos funcionales astrocíticos, neurociencia.
Abstract
Astrocytes are the most abundant cells in the human central nervous system (CNS). Traditionally, astrocyte-related research has been built under the assumption of two basic premises, that astrocytes form a homogeneous syncytium throughout the CNS, and that they have merely supportive and homeostatic functions. The rupture with these premises has led to a new paradigm in astrocyte-related research where a functional network of information-processing and behaviour-linked astrocytes become relevant. Using the “astrocytic ensemble” definition of Delgado et al., this study examines these structures all over the central nervous system describing defined astrocytic ensembles and astrocytic subsets that could also be defined as astrocytic ensembles.
Keywords: astrocytes, ensembles, astrocytic ensembles, neuroscience.
1. Introduction
Astrocytes are the most abundant cells in the human central nervous system (CNS). Traditionally, astrocyte-related research has been built under the assumption of two basic premises, that astrocytes form a homogeneous syncytium throughout the CNS, and that they have merely supportive and homeostatic functions. These include ion buffering, neurotransmitter recycling, blood-brain barrier maintenance, cholesterol synthesis, immune signaling, gliotransmitter release, neurotrophin secretion and regulation of neuronal synaptogenesis and elimination among others (1–4).
Over the last decade, research has shown that the role of astrocytes and neuroglia in brain function has been underestimated (5). Consequently, the paradigm of astroglia investigation has shifted into understanding astrocytes as computational elements which are able to store information in dynamic Ca²⁺ states within their processes (6). Ca²⁺ elevations in astrocytes induce release of gliotransmitters, thus stimulating synaptic receptors and acute synaptic modulation among others (7). Recent studies have identified other ways of storing information in astrocytes, such as cAMP levels (8).
On the other hand, the development of genetic and molecular typification techniques has led to the identification of different astrocytic subpopulations. Astrocytes present specific gene programs that indicate some degree of specialization in their functions depending on their region, developmental origin and local neuronal partners (6). Additionally, astrocytes even exhibit physiological differences within the same brain regions (6), which illustrates a degree of complexity that remains to be fully elucidated.
The investigation into the functional properties of astrocytes led to the identification of the work by Williamson et al. (9): “Learning-associated astrocyte ensembles regulate memory recall”. This study raised questions regarding the diversity of astrocytic ensembles that could be described within the CNS, prompting a review of those already documented. The research revealed that, beyond those associated with memory and learning, few precisely defined astrocytic ensembles have been described in the literature to date (9).
Identification of astrocytic ensembles could further clarify their role in brain function and deepen the understanding of astrocyte-related and idiopathic neurological pathologies. Astrocytes are known to participate in the pathogenesis of a wide range of neurological disorders. For instance, control of CNS damage in neurotrauma, stroke, neuroinfection and autoimmune attack; genetic astrocytopathies; epilepsy; migraine; neuropsychiatric disorders; neurodevelopmental disorders; neurodegenerative diseases, among others. Developing our knowledge about astrocytic ensembles is fundamental to achieving a better comprehension of how our mind works (10).
The objective of this review is to be a scientific dissemination over astrocytic ensembles, to summarise the identified astrocytic ensembles and to propose locations in the CNS where new astrocytic ensembles could potentially be defined.
2. Ensemble definition
This review employs the term astrocytic ensembles, which was proposed by Delgado et al. (11). This definition was chosen as Delgado et al. (11) were the first to define an astrocyte subset from a functional point of view. Thereby, astrocytic ensembles are defined as in vivo functional networks, “modules” or “subpopulations” of astrocytes that activate in a specific behavioral paradigm and participate actively in information-processing.
The concept of astrocytic ensembles completely excludes in vitro models, as it gains meaning in physiological context, where individual astrocytes create functional networks that interact with neurons, other glial cells and the CNS environment. However, identifying specific astrocytic ensembles is highly complex due to the lack of available methods to track in vivo astrocyte activity (11).
3. Described ensembles
3.1. Learning and Memory-associated Astrocytic Ensembles
Real time astrocytic activity monitoring techniques have allowed the identification of Learning and Memory-associated Astrocytic Ensembles (9,11–15). By analysing Ca2+ transient and cAMP dynamics astrocyte subsets which activate in fear-conditioned specific contexts in mouse hippocampus can be identified (9,12). Neuron induction of c-Fos expression in these astrocyte subsets during hippocampal activation (9), as well as the required activation of particular α7-nicotinic acetylcholine receptors in neuron-astrocyte signaling (14) show further evidence of the existence of identified astrocytic ensembles associated with neuron engrams in the hippocampus.
Engrams are understood as the physical substrate of memory and learning, that is to say, a subset of neurons which undergo a persistent physical-biochemical change and synapse-strengthening following a specific experience or event. Consequently, exposure to similar experiences in the future will produce engram activation, leading to what is known as memory recall (16).
Williamson et al. (9) prove the existence of Learning-associated Astrocyte Ensembles (LAA) in a series of consecutive experiments in mice. By analysing the differential expression of c-Fos in different astrocytes in the hippocampus, a subset of astrocytes which underwent experience-dependent activation was found. Selective inhibition of c-Fos expression in those astrocytes significantly worsened hippocampal function. This was shown by diminished freezing behaviour after fear conditioning. An increase in Ca2+ metabolism was also detected. Thus, only LAA ensembles expressed c-Fos and an increase in Ca2+ metabolism because of experience. A specific protein expression, nuclear factor I-A, was also found among LAA ensembles. Moreover, engram-engram synapses were found to be enriched by LAA processes which regulated synaptic plasticity and Long Term Potentiation. Astrocytic ensemble activation was proved to be essential for memory regulation, specifically memory recall (9).
Dewa et al. (12) show how astrocytic ensembles stabilize memory through influencing neuron ensembles and allow recall. Specific subsets of mice hippocampal astrocytes not only undergo experience-dependent activation but also are capable of information processing and integration through noradrenaline-dependent mechanisms. Astrocytic ensembles can link discrete fear experiences at a different pace than engram neurons and may partially meet the criteria for engram cells that undergo physical-biochemical changes upon experiences. However, astrocytic function is rather linked to memory stabilisation and recall than to memory encoding itself (12).
The importance of finding specific activation mechanisms of Learning and Memory-associated Astrocytic Ensembles should be remarked, as the activation of certain pathways does not alter engram function in any way. Astrocytic ensemble roles in the hippocampus may not be restrained to memory stabilisation and recall neither (17).
Romero et al. (13) suggest that historically the predominant neurocentric approach to memory research has excluded non-neuron cells such as astrocytes from participating in brain function, resulting in a lack of understanding memory and learning. Thus, the term Astroengram is proposed to describe how learning experiences activate not only a neuron engram but also associated astrocytic ensembles. The activation of both associated structures will be necessary for memory stabilisation and recall (13).
3.2. Reward Circuit-associated Astrocytic Ensembles
Another identified astrocytic ensemble is found in the reward circuit (mesocorticolimbic pathway), specifically in the nucleus accumbens (NAc).
It is known that astrocytes modulate neuronal activity influenced by tetrahydrocannabinol (THC) through cannabinoid receptors type 1 (CB1R)-induced Ca2+ elevations, which activates presynaptic metabotropic glutamate receptors in tripartite astrocyte-neuron synapses in the mesocorticolimbic pathway. However, whether this mechanism is sufficient to recruit pathway-specific neuron-astrocyte networks has remained unclear (18–20). CB1R expression is non-homogeneous across astrocyte populations. Recent spatial transcriptomic analysis has identified distinct astrocytic subpopulations that completely lack Cnr1 mRNA, such as those in the paraventricular nucleus (19), contrasting with the robust expression found in NAc (20).
By studying THC-induced maladaptive Long Term Depression (LTD) in adolescent mice, Martín-Monteagudo et al. (20) demonstrated that this accumbal astrocytic ensemble is a critical determinant of ventral hippocampus–NAc circuit integrity. It was shown that the aberrant increase in the frequency of astrocytic Ca2+ events and the subsequent p38α MAPK-dependent glutamate release were not merely bystanders, but the actual drivers of the cognitive and behavioral deficits observed. Characterizing anomalies in astrocyte ensembles provides potential therapeutic targets as Martín-Monteagudo et al. described, as the induction of astrocyte ensemble activity is sufficient to restore the cognitive performance of THC-treated mice. These findings suggest that some accumbal astrocytes constitute an ensemble, based on the premise that astrocyte network malfunction leaves neurons unresponsive to synapse stimuli and silences the pathway (20).
4. Possible astrocytic ensembles
Very few astrocytic ensembles have been identified up to date, as there has been little research on this topic. Here, astrocyte networks and subsets that could be identified as astrocytic ensembles with further investigation are reviewed. The focus remains on identifying information-processing behaviour-linked functional properties rather than structural or support properties, following the established definition for astrocytic ensembles.
4.1. Fear-associated Astrocytic Ensembles – Basolateral Amygdala
It has been suggested that a subset of astrocytes in the basolateral amygdala (BLA) participates in the acquisition and maintenance of contextual fear, primarily leading the initial shock response. Precisely, BLA astrocytes are thought to modulate the initial and final motor responses. To correctly predict the outcome of fear conditioning, BLA astrocytes subsets have to actively encode kinematic information and motion related out-put, as shown in Suthard et al. (21) findings. However, BLA astrocytes only influence the fear response in contextual recall, and not in first time responses. These findings raise questions about whether BLA astrocytes are part of a broader ensemble related to memory recall, as described earlier in this article. BLA astrocytes have been categorized as an independent ensemble according to evidence that shows how they modulate fear responses by implicating negative emotional regulation. Consequently, this possible ensemble is independent from memory and learning-associated ensembles previously described and could be part of a reward system astrocyte ensemble that stores aversive experience information (21).
4.2. Circadian rhythms-associated Astrocytic Ensembles – Suprachiasmatic nucleus
In parallel to the BLA findings, the suprachiasmatic nucleus harbors a distinct population of astrocytes that modulate the transcriptional/post-translational feedback loop responsible of the master circadian clock of mammals. Astrocytes in the suprachiasmatic nucleus can maintain and initiate circadian rhythms by robust molecular oscillations in absence of neuron activity (22).
Findings by Patton et al. (23) demonstrate that astrocytes can regulate steady-state CNS circadian oscillations and initiate de novo rhythms with a fidelity comparable to that of neurons. Molecular analyses utilizing Gi- and Gq-DREADDs reveal distinct intracellular pathways. Notably, while the impairment of these pathways disrupts circadian oscillations, it does not entirely inhibit rhythmic activity. This suggests that while astrocytes are necessary for homeostatic precision, they may not be sufficient to independently reset the ensemble phase. As observed in other regions, astrocytes exhibit slower kinetics – measured by the time required for the stabilization of circadian rhythms – compared to neurons (23). This may imply that kinetic dissociation is ubiquitous across the CNS.
Hastings et al. (24) demonstrated that while the CNS can compensate for arginine vasopressin neuronal disruption, the excision of astrocytic Bmal1 induces a foundational alteration that bypasses these compensatory mechanisms. This suggests that astrocytes are not merely contributors, but non-redundant regulators of the circadian ensemble. While neurons remain indispensable for circuit execution, astrocytic failure triggers a primary instability that the network cannot overcome, which proves their role as the obligatory scaffold for rhythm maintenance (24).
4.3. Movement-associated Astrocytic Ensembles – Motor cortex
Delepine et al. (25) findings suggest that M1 cortical astrocytes regulate learning-associated stereotyped movements and dexterity in motor learning through glutamate release. Thus, a new possible astrocytic ensemble could be identified (25).
4.4. Pain-associated Astrocytic Ensembles – Spinal Cord
The requirement for coordinated astrocytic activity is not unique to the brain, as neuron-dependent astrocytic activity has been described in the spinal dorsal horn (26). Astrocytes actively create calcium signals in response to high amplitude noxious stimuli, although the mechanisms underlying these interactions remain unknown (26,27). These could lead to the identification of a new possible astrocytic ensemble, since large scale experience-sensible Ca2+ dynamics is found. Ca2+ transients encode the frequency and intensity of the stimulus and functionally recruit adjacent non-stimulated astrocytes from the ipsilateral and contralateral dorsal horns and from the rostral medullary segments (27). This assertion should be viewed with caution, as the evidence comes from studies evaluating mechanical stimulation (i.e., nociceptive and non-nociceptive stimuli) and studies that do not examine intersubject variability in pain perception (27).
5. Neurogliovascular ensembles
The notion that astrocytic networks behave as a functional syncytium due to being linked by gap junctions appears in the 1970s thanks to the work of Kuffler et al. and has been extensively reviewed (28). The organization of these glial cells in cortical columns and its participation in numerous functions such as energy supply, neural function regulation, blood flow control and protection (in the blood-brain barrier) among others (28), lead to the idea of the existence of NeuroGlioVascular Ensembles (NGVE) (5). These are functional and structural cortical subsets which include astrocytes, neurons and vascular elements.
This concept combines both the ideas of astrocytes being relevant in support functions and information processing at the same time. The threshold between what is just structural, metabolic, and functional support and what can influence brain functions such as information processing and behaviour could be a very fine line.
Specialisation of NGVE along the central nervous system is also possible, as observed in the glomeruli of the olfactory bulb. These differences can be manifested both at a structural and molecular level. Specific connexins may act as biomarkers for identification of different types of NGVE (29)
6. Strengths and limitations.
This main strength of this article is that the definition of an “astrocytic ensemble” was rigorously identified during the literature search. The review which coined the term is recent and uses literature from the last ten years. It belongs to a long-lasting research line about astrocytes by Delgado et al. (11).
Some limitations were also noted during this study. This research’s main limitation is inherent to a narrative review, given the lack of a search protocol. A subsequent systematic review could address this bias. Publication bias has not been controlled as gray literature was not included. Lastly, the reviewed articles used different methods to identify astrocytic subsets, relying on their own, non-standardized procedures that primarily measure Ca²⁺ transients. A lack of available in vivo astrocyte activity tracking methods has been noted.
7. Conclusions
To date, only two astrocytic subsets have been defined as astrocytic ensembles. These are learning and memory-associated astrocytic ensembles in the hippocampus and reward circuit-associated astrocytic ensembles in the mesocortical pathway, specifically in the NAc.
However, the analysis of multiple astrocytic subsets following information-processing behaviour-linked functional properties led to the identification of multiple possible astrocytic ensembles in different parts of the CNS. Basolateral amygdala, motor cortex, suprachiasmatic nucleus and even the spinal cord are some of the places where astrocytic ensembles could potentially be found. Moreover, participation of both neuronal and vascular elements in astrocytic ensembles could result in wider concepts with more profound functional implications. (Figure 1 illustrates the different possible and identified astrocytic ensembles reviewed).
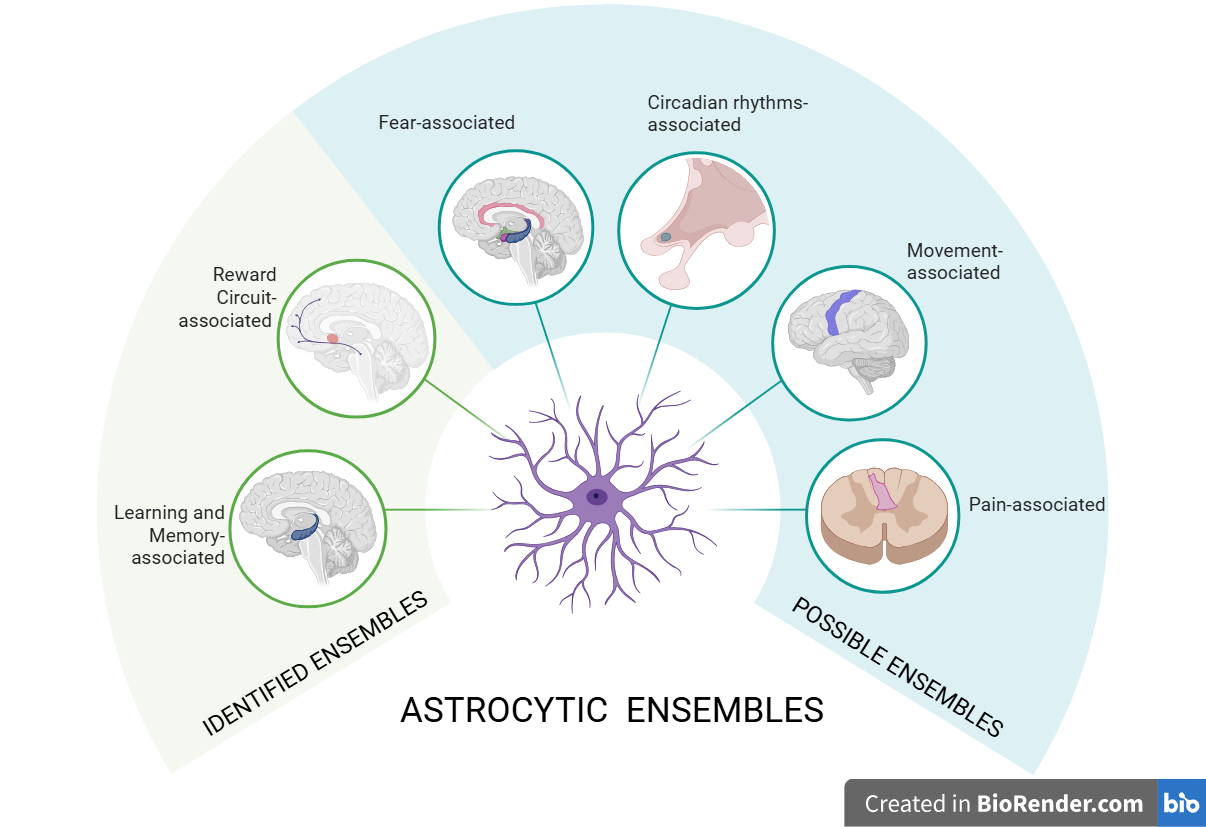
Therefore, astrocytic ensembles are in vivo functional astrocytic networks that activate in specific behavioral paradigm and participate actively in information-processing. The identification of various defined astrocytic ensembles as well as astrocytic network which behave ensemble-like makes it plausible for astrocytic ensembles to exist all over our CNS
Further research is needed to identify and classify these possible ensembles as proper astrocytic ensembles as well as to find more new astrocytic ensembles. Future investigations could not only decipher the way astrocytic ensembles influence our current understanding of the CNS but also their role in CNS pathology.
8. Conflicts of Interest
The authors declare no conflict of interest.
9. Fundings
None
References
1. Miller SJ. Astrocyte Heterogeneity in the Adult Central Nervous System. Front Cell Neurosci. 2018;12:401.
2. Chen PS, Peng GS, Li G, Yang S, Wu X, Wang CC, et al. Valproate protects dopaminergic neurons in midbrain neuron/glia cultures by stimulating the release of neurotrophic factors from astrocytes. Mol Psychiatry. 2006;11(12):1116-25.
3. Phatnani H, Maniatis T. Astrocytes in Neurodegenerative Disease. Cold Spring Harb Perspect Biol. 2015;7(6):a020628.
4. Phatnani HP, Guarnieri P, Friedman BA, Carrasco MA, Muratet M, O’Keeffe S, et al. Intricate interplay between astrocytes and motor neurons in ALS. Proc Natl Acad Sci. 2013;110(8):E756-65.
5. Kirichenko EY, Povilaytite PE, Logvinov AK, Kirichenko YG, Filippova SY. Gap Junctions in the Composition of Neurogliovascular Ensembles in Rat Cortical Barrel Columns. Cell Tissue Biol. 2018;12(6):477-83.
6. Yamamoto M, Takano T. Astrocyte-Mediated Plasticity: Multi-Scale Mechanisms Linking Synaptic Dynamics to Learning and Memory. Cells. 2025;14(24):1936.
7. Makovkin SY, Shkerin IV, Gordleeva SY, Ivanchenko MV. Astrocyte-induced intermittent synchronization of neurons in a minimal network. Chaos Solitons Fractals. 2020;138:109951.
8. Sitja-Roqueta L, Ngum NM, Zherebtsov EA, Kucukerden M, Givehchi M, Bova V, et al. Photoactivated adenylyl cyclase in cortical astrocytes promotes synaptic potentiation and reveals alterations in Huntington’s disease. ISCIENCE. 2025;28(11).
9. Williamson MR, Kwon W, Woo J, Ko Y, Maleki E, Yu K, et al. Learning-associated astrocyte ensembles regulate memory recall. Nature. 2025;637(8045):478-86.
10. Verkhratsky A, Butt A, Li B, Illes P, Zorec R, Semyanov A, et al. Astrocytes in human central nervous system diseases: a frontier for new therapies. Signal Transduct Target Ther. 2023;8(1):396.
11. Delgado L, Navarrete M. Shining the Light on Astrocytic Ensembles. Cells. 2023;12(9).
12. Dewa KI, Kaseda K, Kuwahara A, Kubotera H, Yamasaki A, Awata N, et al. The astrocytic ensemble acts as a multiday trace to stabilize memory. Nature. 2025;648(8092):146-56.
13. Sánchez Romero J, Navarrete M. Astroengrams: rethinking the cellular substrate for memory. Nat Rev Neurosci. 2026;27(4):289-300.
14. Zhang K, Förster R, He W, Liao X, Li J, Yang C, et al. Fear learning induces α7-nicotinic acetylcholine receptor-mediated astrocytic responsiveness that is required for memory persistence. Nat Neurosci. 2021;24(12):1686-98.
15. Holt LM, Nestler EJ, Olsen ML. Cellular and molecular mechanisms of astrocyte plasticity in learning and memory. Trends Neurosci. 2026.
16. Josselyn SA, Köhler S, Frankland PW. Finding the engram. Nat Rev Neurosci. 2015;16(9):521-34.
17. Mak A, Abramian A, Driessens SLW, Boers-Escuder C, Loo RJ van der, Smit AB, et al. Activation of Gs Signaling in Cortical Astrocytes Does Not Influence Formation of a Persistent Contextual Memory Engram. eNeuro. 2024;11(6).
18. Navarrete M, Araque A. Endocannabinoids potentiate synaptic transmission through stimulation of astrocytes. Neuron. 2010;68(1):113-26.
19. Tretiakov EO, Hevesi Z, Böröczky C, Alpár A, Harkany T, Keimpema E. Molecular Fingerprint of Endocannabinoid Signaling in the Developing Paraventricular Nucleus of the Hypothalamus as Revealed by Single-Cell RNA-Seq and In Situ Hybridization. Cells. 2025;14(11).
20. Martin-Monteagudo C, Esparza J, Navarrete M. Astrocytic ensembles in nucleus Accumbens: What role do they play in THC-Related impairments? J Neurochem. 2023;166:22-22.
21. Suthard RL, Senne RA, Buzharsky MD, Pyo AY, Dorst KE, Diep AH, et al. Basolateral Amygdala Astrocytes Are Engaged by the Acquisition and Expression of a Contextual Fear Memory. J Neurosci. 2023;43(27):4997-5013.
22. Brancaccio M, Edwards MD, Patton AP, Smyllie NJ, Chesham JE, Maywood ES, et al. Cell-autonomous clock of astrocytes drives circadian behavior in mammals. Science. 2019;363(6423):187-92.
23. Patton AP, Smyllie NJ, Chesham JE, Hastings MH. Astrocytes Sustain Circadian Oscillation and Bidirectionally Determine Circadian Period, But Do Not Regulate Circadian Phase in the Suprachiasmatic Nucleus. J Neurosci. 2022;42(28):5522-37.
24. Hastings MH, Smyllie NJ, Patton AP. Molecular-genetic manipulation of the suprachiasmatic nucleus circadian clock. J Mol Biol. 2020;432(12):3639–60.
25. Delepine C, Shih J, Li K, Gaudeaux P, Sur M. Differential Effects of Astrocyte Manipulations on Learned Motor Behavior and Neuronal Ensembles in the Motor Cortex. J Neurosci. 2023;43(15):2696-713.
26. Sekiguchi KJ, Shekhtmeyster P, Merten K, Arena A, Cook D, Hoffman E, et al. Imaging large-scale cellular activity in spinal cord of freely behaving mice. Nat Commun. 2016;7:11450.
27. Rivera-Villaseñor A, Higinio-Rodríguez F, López-Hidalgo M. Astrocytes in pain perception: A systems neuroscience approach. Adv Neurobiol. 2024;39:193–212.
28. Giaume C, Koulakoff A, Roux L, Holcman D, Rouach N. Astroglial networks: a step further in neuroglial and gliovascular interactions. Nat Rev Neurosci. 2010;11(2):87-99. doi:10.1038/nrn2757
29. Kirichenko EY, Logvinov AK, Filippova SY, Arefiev RA, Semynina VG, Lysenko LV. Structural Features of Neuro-Glio-Vascular Ensembles in the Glomeruli of the Rat Olfactory Bulb. Cell Tissue Biol. 2020;14(5):372-9.
AMU 2025. Volumen 8, Número 1
Fecha de recepción: 07/02/2026
Fecha de revisión: 28/02/2026
Fecha de aceptación: 10/03/2026
Cita el artículo: Nasi-García, Luca, Jiménez-González, Sonia. Searching for astrocytes: a review on astrocytic ensembles across the central nervous system .AMU. 2026;8(1):20-26.
URI: https://hdl.handle.net/10481/105370